 |
IV.7. fejezet: Molekuláris citogenetikai vizsgálati lehetőségek
A citogenetika egy olyan tudomány terület, amely a kromoszómák szerkezetével, tulajdonságaival, valamint a sejtosztódáskor játszott szerepükkel foglakozik. Nem csak azt vizsgálja, hogy normális esetben hogyan viselkednek a kromoszómák, hanem azt is hogy milyen faktorok indukálnak a kromoszómákban szerkezeti változásokat, milyen jellegű változások ezek, és ezek miként befolyásolják a normális fenotípust. Hagyományos citogenetikai vizsgálati módszernek nevezzük az ún. sávozási technikákat, amellyel a kromoszómák megkülönböztethetővé válnak és a méretük ill. sávmintázatuk alapján áttekinthető a teljes kromoszómakészlet (Ez tehát egy ún. genom szintű vizsgálat. NEM génszintű, és nem egyenlő a laikusok által „géntérképnek” nevezett módszerrel, de kromoszóma-szinten a teljes humán genetikai anyag áttekintésére alkalmas)
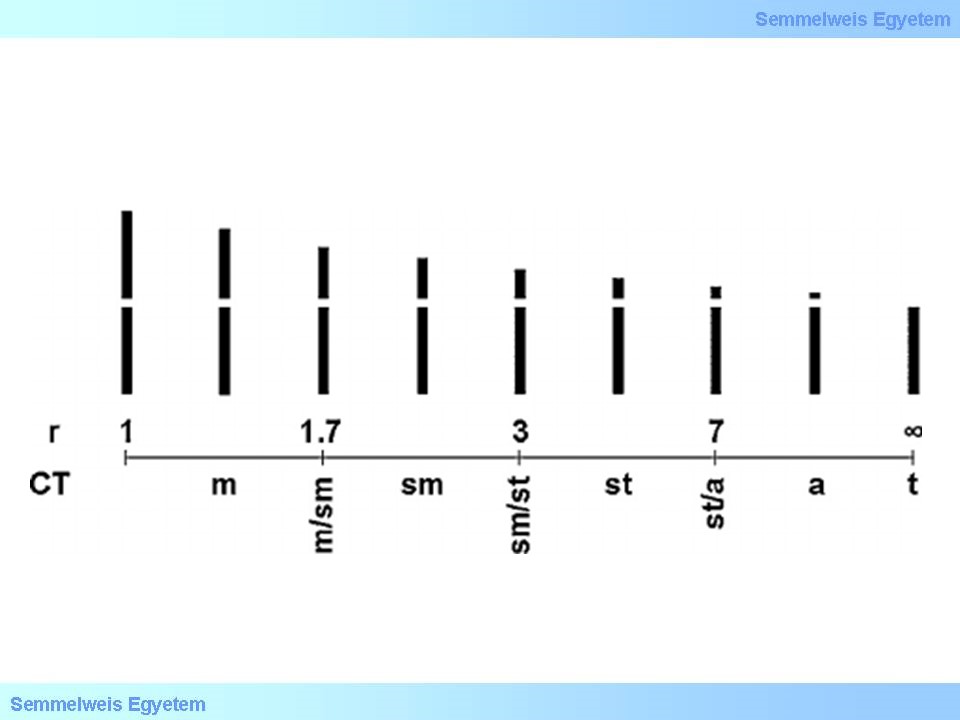
A módszer felbontása 10Mb körüli, tehát legalább 10 millió DNS bázispárnak kell hiányoznia vagy megkettőződnie ahhoz, hogy G-sávozással észrevehető legyen. A hiány általában jobban felfedezhető, mint a többlet. A sávozási technika rutinszerű alkalmazása széles körben elterjedt, egyetlen korlátja az alacsony felbontóképessége. A technikai fejlődésnek köszönhetően megjelentek azok az ún. új molekuláris citogenetikai vizsgálati módszerek (FISH és array CGH technika), amelyek nagyobb felbontóképességgel rendelkeznek a sávozási módszerekhez képest és ezzel lehetővé teszik a kisebb kromoszómális eltérések detektálását is.
|
 |
Mérföldkövek
A klasszikus citogenetikai vizsgálatoktól az új molekuláris módszerekig.
-
- J. Arnold: az első kromoszóma megfigyelése (1879) (22. ábra).
-
- W. Flemming: elsőként írta le a kromoszómákat és viselkedésüket a mitózis során (1882) (23. ábra).
-
- Jo Hin Tijo és A. Levan: leírták a pontos emberi kromoszóma számot 46 (The chromosome number of man. Hereditas 1956; vol. 42, pages 1–6.) (24. ábra.).
-
- Lejeune: a 21. kromoszóma triszómiáját Down szindrómának nevezte el (1959).
-
- Levan: kromoszómák osztályozásának bevezetése (1964).
-
- M.Drets és M. Shaw: G-sávozás bevezetése (1971).
-
- Az első nemzetközi humán citogenetikai nevezéktan megjelenése (1995)

22. ábra: Az első rajzolt kromoszóma kép J. Arnold heidelbergi patológus által megjelentett publikációból az “Archiv für Pathologische Anatomie” újságban 1879 © University Hospital Heidelberg.
|

23. ábra: Zellsubstanz, Kern und Zelltheilung, 1882 W. Flemming és az általa rajzolt sejtek a mitózis alatt.
|
24. ábra: A. Levan, K. Fredga, A. A. Sandberg. Nomenclature for centromeric position on chromosomes. Hereditas:52, 2, 1964; 201–220.
|
A számbeli kromoszómaeltérések (aneuploidiák - extra vagy hiányzó kromoszómák, számfeletti marker kromoszómák) meglehetősen ritkák, a postnatalis élettel akár csak rövid ideig is csupán a 13-as, 18-as, és 21-es autoszómák többlete egyeztethető össze, és egyetlen autoszóma hiánya sem. Az X és Y kromoszómák, vagyis a gonoszómák többlete vagy hiánya lehetővé teszi a méhen kívüli életet. Astrukturális kromoszómális eltérések, mint pl. deléciók, duplikációk transzlokációk, inverziók aneusomiát, azaz az átrendeződésben részt vevő kisebb kromoszómaszakaszok elvesztését vagy többletét okozhatják.
Minden olyan állapotban, amely látható veleszületett rendellenességgel és/vagy értelmi elmaradással jár, lányok nem tisztázott eredetű alacsonynövése, továbbá a nemi érés zavaraiban, meddőség vagy ismétlődő vetélések esetén érdemes elvégezni a kariotipizálást.
Az értelmi elmaradás (mentális retardáció) előfordulási gyakorisága az átlagpopulációban megközelítőleg 3%, a veleszületett rendellenességeké kb.1%-ra, az autizmus spektrum zavaré (ASD) 1:150-re tehető.
Ezen állapotok hátterében mintegy 6%-ba azonosítható kromoszomális ok. Amennyiben G-sávozással nem mutatható A FISH technikát alkalmazva, a submikroszkópikus és subtelomerikus kiegyensúlyozatlan eltéréssel járó ismeretlen eredetű mentális retardációs kórképek 2.5-7%-ban azonosíthatók. A szintén genom szintű array komparatív genomiális hibridizáció (arrayCGH) alkalmazásával 15-20%-ra növelhető a diagnosztikus arányt.
A FISH felbontóképessége 3-5 Mb, az array CGH-é jellemzően 12-20 Kb. Míg a FISH kizárólag célzott régiók vizsgálatára alkalmas, az array CGH a kariotipizáláshoz hasonlóan genom szintű vizsgálat, de sokkal jobb felbontással. Szülői kiegyensúlyozott transzlokációk kimutatására azonban az array CGH nem alkalmas, a FISH és a kariotipizálás igen.
|
 |
Eset ismertetés
Klinikai tünetek
Egy fiatal Turner szindrómával diagnosztizált női beteg molekuláris citogenetikai vizsgálatát mutatjuk be. A beteg átlagnál alacsonyabb és vékonyabb termetű (ösztradiol terápiában részesült ezért a másodlagos nemi jellegek kifejlődtek, tíz éves kora óta halláskárosodott és gyakori vesemedence-gyulladásai vannak. Tizenöt éves korában diagnosztizálták cukorbetegségét és súlyos vérszegénységét, valamint lisztérzékenységét. Nőgyógyászati vizsgálat 23 éves korában történt és mindkét petefészeknél a normálistól kisebb méretet állapítottak meg.
Molekuláris citogenetikai vizsgálatok
G sávozás perifériás lymphocita sejtekből, FISH vizsgálat X és Y centromer próbákkal, valamint teljes X és Y festő próbákkal, további specifikus FISH festés X p kar piros, Xq kar zöld, (SHOX, XIST) és subtelomer Xq/Yq specifikus próbákkal történt a töréspontok azonosítása céljából. Az array-CGH vizsgálatot ISCA plus NimbleGene-Roche rendszerrel végeztük. A specifikus FISH próba és array CGH eredmények alapján egy normál X kromoszóma és egy többszörösen eltört és átrendeződött X kromoszómát azonosítottunk. Az array CGH eredmények a 25. ábrán láthatóak. A molekuláris citogenetikai vizsgálati panel a 26. ábrán szerepel.

25. ábra
|
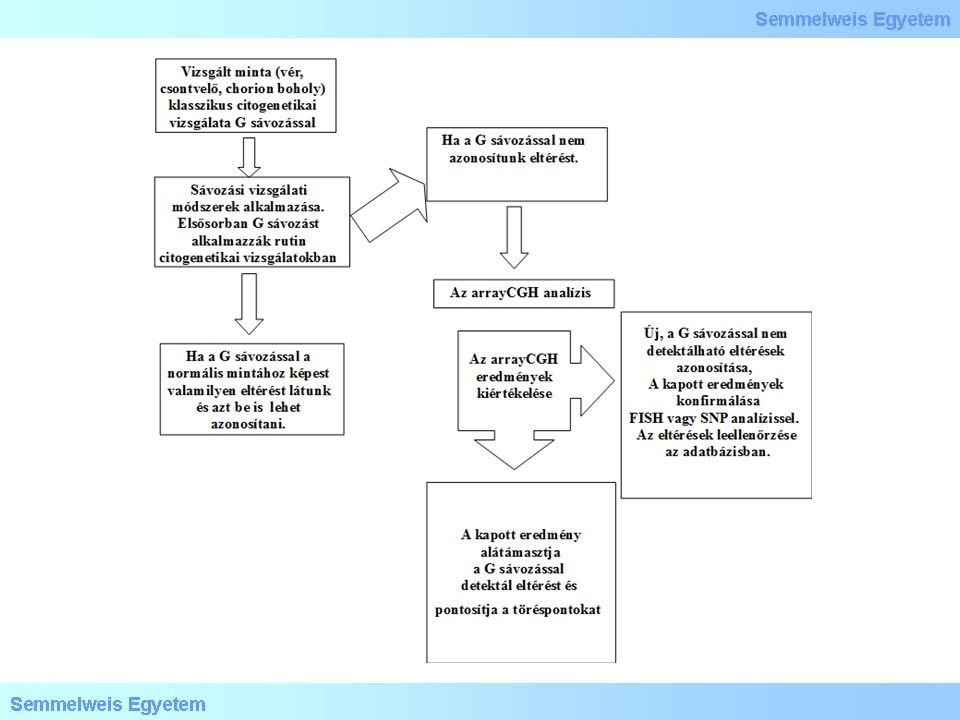
26. ábra: Molekuláris citogenetikai vizsgálati panel
|
Eredmények
Array-CGH (NimbleGene Array CGX 1.4 M; assembly GRCh37/hg19) egy kópiát detektál az X kromoszóma rövid karjából ezért feltételeztük, hogy a másik X kromoszómáról hiányzik a rövid kar (Xp11.1-22.33 ; 60,697 - 58,554,898 ± 58,566,094 bp régió). A hosszú kar Xq 27.3 régiója (142,391,424 ± 142,402,005 – 144,097,010 ± 144,108,359 bp) normális két kópiában volt jelen. Két különböző méretű szegmenst azonosítottunk a Xq karon, amely három kópiában volt jelen: 80,3 Mb nagyságút az Xq11.1 - Xq27.3 régióban és a másik egy kisebb méretűt (11,15Mb) az Xq27.3 - Xq28 régióban. Az array-CGH eredmények, valamint a Specifikus FISH próbák alapján kiraktuk a beteg X kromoszómájának virtuális kariogramját.
|
|